Молекули імунної системи
ІМУНОГЛОБУЛІНИ
Молекули імуноглобуліну (Ig) — це ефекторні продукти В-клітин і, незважаючи на те, що всі вони дуже схожі за своєю будовою, незначні розбіжності в межах головних клясів (IgG, IgA, IgM, IgD та IgE) та підклясів (IgG 1, 2, З та 4; IgA 1 та 2) пов'язані з рядом важливих біологічних властивостей (табл. 2.3). Молекули, майже тотожні виділеним імуноглобулінам, накопичуються у клітинних мембранах В-клітин (поверхневі Ig). Існує велика кількість зв'язаних з ними молекул, які беруть участь у розпізнаванні антигенів та у процесі зв'язку клітин.
Будова молекул імуноглобуліну
Молекула імуноглобуліну складається з чітко визначених субодиниць, поєднаних між собою за допомогою дисульфідних зв'язків (S — S), які руйнуються під дією послаблюючих фак¬торів, і молекула розривається на пари поліпептидних ланцюгів, що називаються важкі та легкі ланцюги. Існують два типи легких ланцюгів, капа та лямбда, з них індивідуяльні імуноглобуліни мають тільки один тип, а також кілька типів важких ланцюгів, що надають Ig-молекулі її властивости відповідно до кляси, до якої вона належить.
Звичайна молекула імуноглобуліну, наприклад, IgG (мал. 2.3), має дві ділянки, де зв'язується антиген (Fab), та Fc-компонент, що є частиною молекули, яка виконує функцію зв'язування клясів, як, наприклад, фіксації комплементу. Фраґмент важкого ланцюга, який міститься у Fc-компоненті, відповідає за антигенні відмінності між клясами імуноглобуліну, і це дає змогу визначати їх за лабораторних умов шляхом застосування важколанцюгової спеціяльної антисироватки.
Імуноглобуліни відрізняються за антиген-зв'язуючою функцією, тобто за активністю антитіл, молекулярна основа яких розміщена у так званих V-сеґментах Fab-компонентів. Різноманітні імунокомпетентні клітини здатні розпізнавати та реаґувати на клітинний рецептор антигена молекули імуноглобуліну (ідіотип), виділяючи при цьому антиідіотипові антитіла, що можуть впливати на об'єм та тривалість утворення антитіл для даного антигена.
Імуноглобулін G (IgG)
У здорових дорослих людей більш як 70 відсотків загального вмісту імуноглобулінів припадає на основний сироватковий IgG, він рівномірно розповсюджується між кров'ю та зовнішньоклітинними рідинами. Щодоби майже чверть усієї кількости IgG, що знаходиться в організмі людини, виходить з кровопотоку, і така сама кількість повертається через грудну лімфатичну протоку. IgG — це єдиний у людини імуноглобулін, який проходить через плаценту, доходить до плоду та забезпечує новонароджену дитину пасивно надбаним антитілом, яке функціонуватиме протягом початкового періоду її життя.
IgG-антитіла відіграють дуже важливу ролю у протибактерійному імунітеті. Вони легко нейтралізують розчинні токсини, що є відповідальними за клінічні ознаки дифтерії та правцю. Крім того, IgG здатні опсонізувати бактерії, вони вкривають їх, таким чином полегшуючи поглинання бактерій фагоцитами. Але IgG-антитіла здатні також призводити до розвитку захворювання, наприклад, як аутоантитіла або внаслідок формування імунних комплексів.
Імуноглобулін M(IgM)
Макромолекулярний IgM — це здебільшого внутрішньосудинна речовина. Вона складається з п'яти імуноглобулінових одиниць, що з'єднуються дисульфід ними зв'язками для того, щоб сформувати десять однакових клітинних рецепторів антигена разом із з'єднувальним пептидом (j-ланцюгом). Дія IgM особливо ефективна у процесі активації комплементу задля імунного знищення сторонніх клітин. Крім того, антитіла IgM значно ефективніше, ніж антитіла IgG з'єднують відповідні антигени між собою з метою аґ лютинації та фагоцитозу, а також вони дуже активні при знищенні та нейтралізації продуктів розпаду клітин або бактерій, що перебувають у кровопотоці.
Імуноглобулін A (IgA)
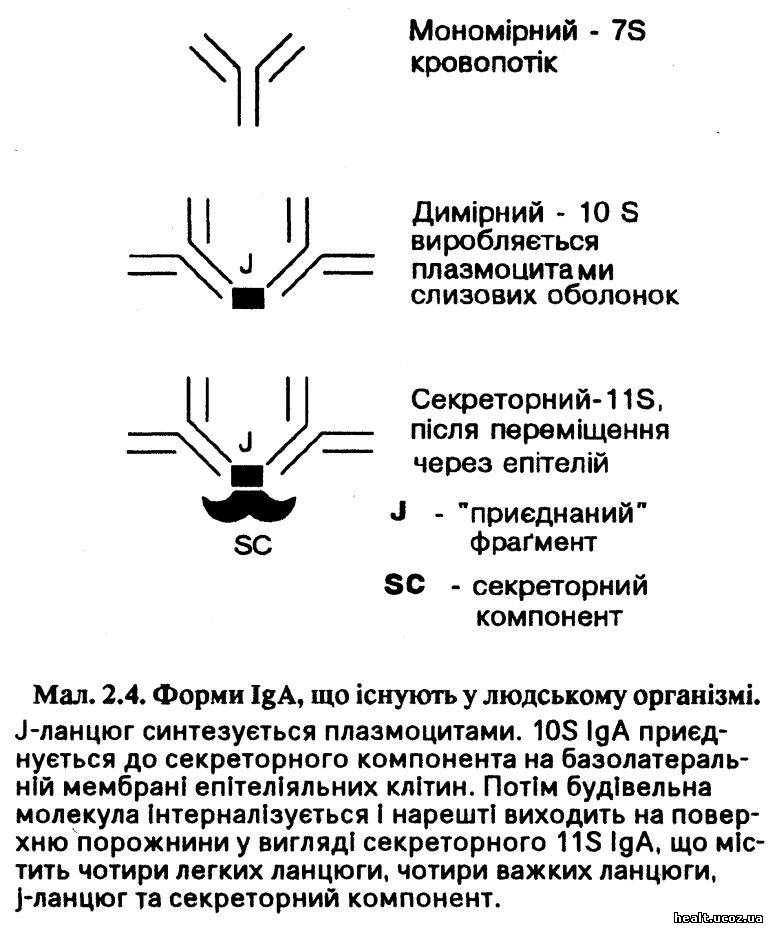
На IgA припадає майже 20 відсотків сумарного вмісту імуноглобулінів у сироватці. Проте вважають, що він виконує незначну або взагалі ніякої функції у межах кровопотоку і тка¬нин, однак його роля як секреторного антитіла значна. Найважливішими зонами синтези IgA є слизові оболонки дихальних шляхів, кишок та інші слизової оболонки. IgA існують у трьох головних молекулярних формах (мал. 2.4). У крові — це 7S, мономірний, за розмірами такий, як IgG. Плазмоцити слизових оболонок виробляють димірний 10S IgA (який також містить j-ланцюг). Через епітелії він потрапляє до молозива, слини, кишкового соку, дихальних секретів, сліз та деяких інших рідин організму. Протягом трансепітеліяльного пересування до нього приєднуються інші поліпептиди, секреторні компоненти, внаслідок чого формується секреторний US IgA, який має відносну опірність щодо ферментів травлення.
Секреторний IgA створює імунітет до кишкових інфекцій та вірусів, а також може брати участь у реґуляції мікрофльори кишечника. У наш час для стимулювання захисного імунітету до кишкових інфекцій, таких як холера та ротавірус, застосовують методу пероральної імунізації.
Імуноглобулін D(IgD)
Майже весь IgD міститься на поверхні незрілих В-лімфоцитів, він може брати участь у процесі їх визрівання та реґуляції.
Імуноглобулін Е (IgE)
Вміст IgE у сироватці дуже низький. Це частково наслідок того, що він значною мірою притягується до поверхні клітин і міцно приєднується до мастоцитів та базофілів. IgE відіграє дуже важливу ролю у реакціях гіперчутливости миттєвого типу, таких, які властиві схильним до алерґії людям, наприклад, при полінозах. Фізіологічна функція IgE-антитіл повністю ще не вивчена, але вже встановлено, що вони відіграють важливу ролю у захисті організму від гельмінтів.
КОМПЛЕМЕНТ
Система комплементу — це розширений каскад, схожий на той, що відповідає за процес згортання крови та фібриноліз. Активація цього ефекторного механізму відбувається або шляхом імунної реакції, опосередкованої антитілами (клясичний шлях), або під дією бактерійних та інших хемічних стимуляторів (альтернативний шлях). Обидва шляхи ведуть до остаточної узгоджености послідовних дій, кульмінацією яких є утворення "мембрано-атакуючого комплексу", де відбувається формування канальців, які пересікають клітинну мембрану, й це призводить до знищення клітин.
Компоненти комплементу
Існує узгоджена термінологія, яку застосовують для позначення компонентів, наприклад, С2, СЗ, але вони функціонують не відповідно до своїх ідентифікаційних номерів. Повний перелік компонентів комплементу у будь-якому підручнику з імунології, їхній опис ґрунтується на загальних принципах реакцій та їхньому клінічному значенні. Більшу частину компонентів комплементу скла-дають білки (бета-глобуліни), які утворюються у печінці і після активації виконують функцію протеїнази. Винятком є Clq, який являє собою своєрідну молекулу, конфігурація якої схожа на пучок з шести тюльпанів. Інґібітори також беруть участь у різноманітних діях каскадів та у циклах зворотного зв'язку.
Послідовність дій при активації комплементу
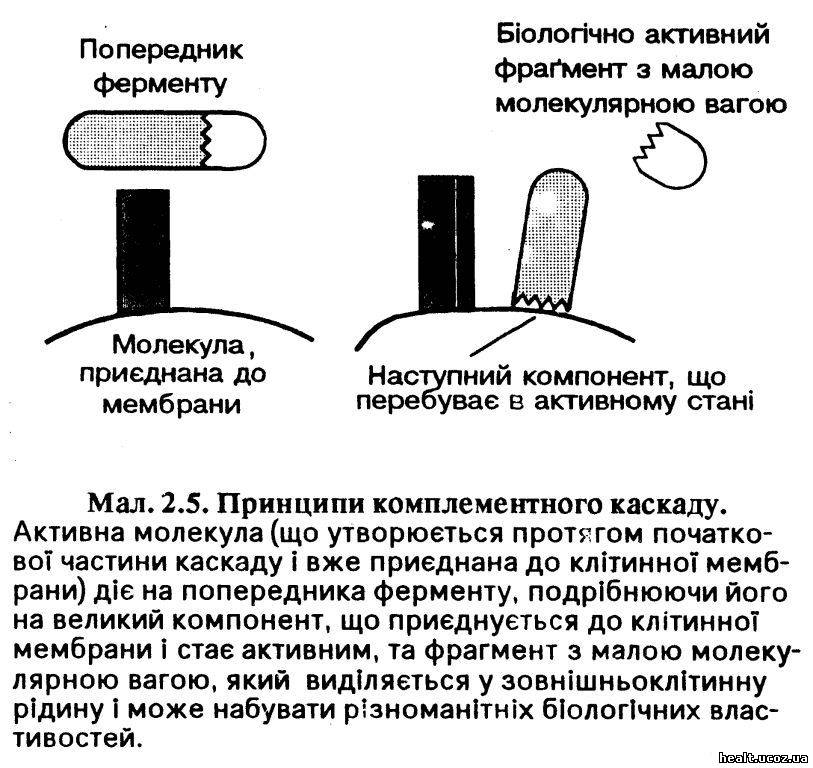
Комплемент включається у склад мембрани або імунного комплексу, де відбувається послідовна активація ферментів комплементу (мал. 2.5). Завдяки дії мембрани та секреції активних пептидів система комплементу виконує ряд функцій, у тому числі активацію клітин, знищення клітин, запалення та опсонізацію.
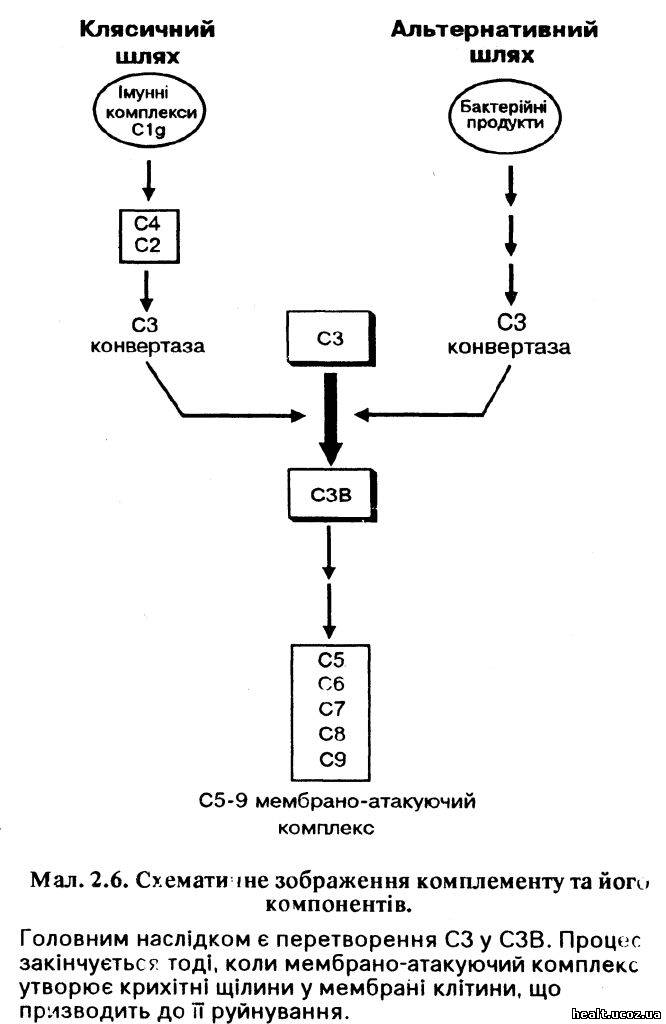
На мал. 2.6 зображено важливі стадії активації комплементу. Активація СЗ за допомогою ферменту — СЗ-конвертази — це центральна подія комплементної послідовносте. Активізація комплементу клясичним шляхом відбувається тоді, коли імунні комплекси об'єднуються з Clq. Коли відбувається активація комплементу альтернативним шляхом, СЗ-конвертаза утворюється внаслідок процесу, що починається з активації компонентів комплементу під дією таких матеріялів, як стінки бактерійних клітин та ендотоксин. Таким чином, альтернативний шлях особливо придатний тоді, коли організм ще не підготував первинної імунної відповіди.
Деякі важливі біологічні функції комплементу наведено у табл. 2.4. Якщо у процесі захворювання бере участь комплементна послідовність, СЗ можна використовувати у великих кількостях. Тому важливе значення для встановлення діагнози має визначення низьких рівнів вмісту цього білка у крові.
Шлях проходження активації комплементу контролюється двома механізмами. Значна кількість активованих компонентів успадковують здатність видозмінюватися. Якщо наступний протеїн каскаду відсутній, то активна речовина розпадається. Існує також ряд інгібіторів, наприклад, інгібітор СІ.
ЦИТОКІНИ
Деякі види цитокінів часто ще називають лімфокінами, інші перейменували в інтерлейкіни.
Наприкінці 60-х років виявили, що біологічно активні розчинні речовини іноді здатні викликати ефекти, опосередковані лімфоцитами. Було описано багато таких речовин. Провели велику кількість досліджень специфічного антигенного клітино залежного імунітету, які ґрунтувались на припущеннях стосовно секреції таких речовин Т-клітинами. Яскравим прикладом є реакція гальмування переміщення лейкоцитів. Зараз ґрунтовно вивчили багато продуктів, що їх виділяють Т-та інші клітини. Визначили їх біохімію та навіть гени, що їх кодують. Стало зрозумілим, що тільки незначна кількість молекул має властивості лімфокінів. Найважливішими є інтерлейкін I (ILI), який виділяють макрофаги; IL2, який виробляють Т-клітини та інтерферони. Більш детальна інформація про них та інші цитокіни наведена у табл. 2.5